| Science | Diverses | Kinderseiten |
| |
Die etwas andere Homepage ... | ||
|
|
|
Topics:
Cell cycle control Der mitotische Zellzyklus wird in vier Abschnitte, die G1-, S-, G2- und M-Phase unterteilt. In der S- und M-Phase erfolgen die beiden grundlegenden Prozesse der Zellproliferation, die DNA-Synthese (Replikation) und die Kernteilung (Mitose). In den dazwischenliegenden Phasen laufen biochemisch schwerer zu charakterisierende, vorbereitende Prozesse ab; daher wurden diese Zeiträume gap-Phasen (engl. Lücke, Pause) genannt (zur Geschichte der Zellzykluserforschung siehe Masui, 1992; Dorée und Galas, 1994). Bereits 1978 postulierte Hartwell, daß der eukaryotische Zellzyklus ein abhängiger Zellzyklus sei, indem verschiedene Phaseneintritte nur dann erfolgen, wenn die vorherigen Prozesse vollständig und korrekt abgeschlossen sind. Dieses Konzept wurde später durch zahlreiche Studien bestätigt, die zeigen konnten, daß alle eukaryotischen Zellen durch externe und interne Einflüsse an mehreren Punkten im Zellzyklus arretiert werden können. Diese Arretierungspunkte werden heute Kontrollpunkte genannt und mit den verantwortlichen Zellzyklusregulatoren im Kontrollpunktmodell zusammengefaßt (Peeper et al., 1994; Elledge, 1996). Vom
Beginn bis in die späte G1-Phase ist das Voranschreiten des Zellzyklus
(Zellzyklusprogression) vom Vorhandensein extrazellulärer Wachstumsfaktoren
und (bei nicht hematopoietischen Zellen) Zell-Zell- bzw. Zell-Matrix-Kontakten
abhängig. Ist einer oder mehrere dieser Faktoren nicht gegeben, so wird
die Zellzyklusprogression blockiert (G1-Arrest oder G1/S-Blockade). Längerer
Entzug von Wachstumsfaktoren führt zum reversiblen Austritt aus dem Teilungszyklus
und dem Eintritt in die G0-Phase oder Quieszenz. Diese ist durch die Expression
sogenannter Markerproteine wie Statin und Gas-Proteine (growth
arrest specific) gekennzeichnet (Gustincich und Schneider, 1993; Sandig
et al., 1994). Hiervon zu Unterscheiden sind die nicht reversible terminale
Differenzierung und die Seneszenz (Zellalterung) (Müller et al., 1993).
Mit dem Überschreiten des R-Punkts (restriction
point) sind diese Faktoren nicht mehr limitierend und der S-Phaseeintritt
erfolgt dann auch unter Serummangelbedingungen (Planas-Silva und Weinberg,
1997).
Abb.:
Zellzyklusphasen und Kontrollpunkte
eukaryotischer Zellen Bis
zur Vollendung der S-Phase detektiert der S/G2-Kontrollpunkt nicht replizierte
DNA und verhindert den Eintritt in die G2-Phase. Ebenfalls in der S-Phase,
jedoch auch weit über diese hinaus wirkt die Endoreduplikationskontrolle,
die gewährleistet, daß jeder DNA-Abschnitt nur einmal pro Zyklus verdoppelt
wird (Nurse, 1994; Grafi, 1998). Am G2/M-Übergang ist der sogenannte NEB
(nuclear envelop breakdown)-Kontrollpunkt lokalisiert. Zellen, die
an diesem Punkt arretiert werden, haben eine intakte Kernhülle und dekondensiertes
Chromatin (King et al., 1994; Rieder und Khojakov, 1997). Der vierte Kontrollpunkt
überprüft in der M-Phase die Spindelbildung und Mikrotubulibindung an
die Kinetochore der Chromosomen und wird daher entweder als Spindelaufbau
(spindle assembly)- oder Kinetochorkontrollpunkt
bezeichnet (King et al., 1996; Rieder und Khodjakov, 1997).
Zellzyklusregulation durch Zyklin-abhängige Kinasen Die grundlegenden Effektoren der Zellzyklusprogression sowie ihrer zeitlichen Koordination und Kontrolle sind die Zyklin-abhängigen Kinasen (cyclin-dependent kinases, Cdk). Diese Kinasen sind als Monomere inaktiv, und ihre katalytische Funktion ist vollkommen abhängig von der Assoziation mit regulatorischen Untereinheiten, den sogenannten Zyklinen. In Hefezellen ist eine einzige Cdk (Cdc2 in Schizosaccharomyces pombe und Cdc28 in Saccharomyces cerevisiae) durch die Assoziation mit bis zu 9 verschiedenen Zyklinen in der Lage alle Phasenübergänge zu kontrollieren (Nigg, 1995; Morgan, 1995). In Säugerzellen dagegen wurden bisher aufgrund von Sequenzhomologien mehr als 10 Cdc2-verwandte Proteine und 13 Zykline identifiziert (Nigg, 1995; Pines, 1995; Morgan, 1997). Die Cdk-Formen der Vertebraten werden durch Numerierung von 1 bis 8 unterschieden, wobei die Cdk1 das Homolog der Cdc2 bzw. Cdc28 aus Hefezellen ist. Alle Cdk-Formen sind 33 bis 40 kDa groß, weisen 35 bis 65% Sequenzhomologie auf und besitzen eine im N-terminalen Lappen lokalisierte, hochkonservative Helix, die für die Zyklinbindung essentiell ist. Diese wird nach der ursprünglichen Aminosäuresequenz in der Cdc2 als PSTAIRE-Helix bezeichnet (Morgan, 1997; Noble et al., 1997). Die
Zykline aus Vertebratenzellen werden alphanumerisch von A bis H benannt.
Innerhalb der Zyklin-Familien A, B und D sind mehrere Formen bekannt,
die nicht durch alternatives Splicen aus einem Gen entstehen. Sie werden
als eigenständige Zykline A1, A2, B1 bis 3 und D1 bis 3 bezeichnet, während
die Splicevarianten z.B. von Zyklin E durch den tiefgestellten Zusatz
s (small)
und l (large)
gegeneinander abgegrenzt werden (Sewing et al., 1994; Nigg, 1995; Pines,
1995). Die Zykline sind eine erstaunlich diverse Familie von Proteinen.
Ihre Größe reicht von 35 bis 90 kDa. Die entfernteren Verwandten zeigen
kaum Sequenzhomologien außer in einer ca. 100 Aminosäuren umfassenden
Region, der sogenannten Zyklinbox, die für die Cdk-Bindung und -Aktivierung
verantwortlich ist (Morgan, 1997; Noble et al., 1997). Ihren Namen erhielten
die Zykline aufgrund ihrer starken Expressionschwankungen im Verlauf des
Zellzyklus, die zumindest zu einem großen Teil die Oszillationen der Cdk-Aktivitäten
begründen (Dorée und Galas, 1994; Nigg, 1995; Pines, 1995; Morgan, 1995). Wie
in Hefezellen binden die zellzyklusregulierenden Cdk-Formen der Vertebraten
meist mehrere Zykline. In der folgenden Tabelle sind die Bindungspartner
aller bekannten Cdk-Formen aus Vertebratenzellen zusammengefaßt. Tab. : Bindungspartner der Zyklin-abhängigen Kinasen
p35:
ist ein Protein, das nur in seiner Tertiärstruktur Homologien zu den Zyklinen
aufweist. Ein anderer Bindungspartner der Cdk5 konnte bisher nicht identifiziert
werden. Die Cdk-Bindungspartner der Zykline F und G
wurden bisher nicht ermittelt. Zyklin F scheint jedoch eine Funktion in
der G2-Phase zu haben. Das Zyklin G-Gen ist p53-responsiv. (Nigg, 1995;
Morgan, 1997) Neben
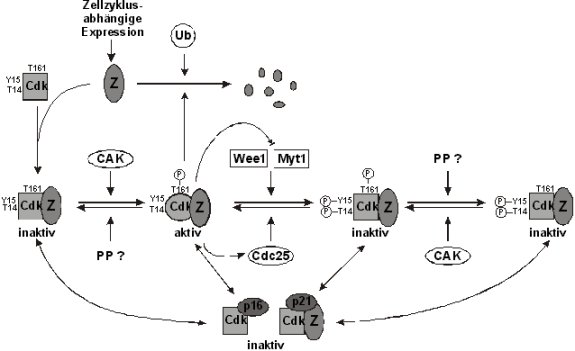
der Zyklinbindung können Cdk-Formen prinzipiell über drei weitere Mechanismen
reguliert werden. Dies sind: I)
die aktivierende Phosphorylierung an einem Threoninrest (T), der
dem T161, der Cdc2 II)
die inaktivierende Phosphorylierung eines Threonin- und Tyrosin
(Y)-Restes im aktiven III)
die Assoziation mit verschiedenen Proteininhibitoren (siehe
Abb., Morgan, 1995; Nigg, 1995).
Abb.:
Prinzipielle Regulationsmechanismen Zyklin-abhängiger Kinasen Der
Ubiquitin-abhängige Abbau der Zykline ist sowohl nach dem S-Phaseeintritt
als auch beim Austritt aus der Mitose am Abschalten der jeweiligen Cdk-Formen
beteiligt. In beiden Fällen sind phasenspezifische Ubiquitin-konjugierende
Enzyme (E2) und Ubiquitin-Ligasen (E3) für die Substraterkennung verantwortlich.
Neben dem Abbau der Zykline D1 und E in der S-Phase bzw. aller Zyklin
A und B-Formen in der M-Phase spielt der Ubiquitin-abhängige Abbau auch
in anderen Zellzyklus-regulierenden Prozessen eine große Rolle. Zu den
bisher identifizierten Substraten gehören u.a. die Cdk-Inhibitoren p21
und p27, E2F-1, E2F-4, p53, IkB
und CUT2/PDS2 (Hochstrasser, 1996; King et al., 1996; Pagano, 1997; Pickart,
1997). Die
Bedeutung der anderen Regulationsmechanismen ist stark phasenabhängig.
Vereinfachend läßt sich sagen, daß die Cdk-Regulation über Bindung an
Proteininhibitoren der Hauptmechanismus der Cdk-Inaktivierung in G1 ist.
Die verantwortlichen Inhibitoren werden in Vertebratenzellen zwei Klassen
zugeordnet. Die Ink4-Familie umfaßt p15-ink4B, p16-ink4A, p18-ink4C und
p19-ink4D. Alle Mitglieder binden freie Cdk4 und 6-Monomere oder verdrängen
die D-Zykline aus der Bindung an diese Kinasen. Die zweite Gruppe umfaßt
p21-waf1/cip1/sdi1, p27-kip1 und p57-kip2. Ihnen gemeinsam ist, daß sie
alle Cdk/Zyklin-Komplexe hemmen und freie Zykline binden (Kato et al.,
1994; Serrano, 1997). In
der S- und G2-Phase dagegen wird der Funktionszustand der Cdk/Zyklin-Komplexe
hauptsächlich über die Phosphorylierung von Threonin 14 und Tyrosin 15
im aktiven Zentrum der Cdk reguliert. In beiden Phasen ist eine basale
Kinaseaktivität nachweisbar, der Großteil zumindest der Cdk1/Zyklin B1-Komplexe
ist jedoch bis zum M-Phaseeintritt inhibitorisch phosphoryliert (King
et al., 1994; Grafi, 1998). Das
Zusammenspiel aller beschriebenen Regulationsmechanismen sowie weiterer
positiver und negativer Rückkopplungsmechanismen (s.u.) resultiert in
einer fein abgestimmten Reihenfolge von Cdk-Aktivierungen und Phaseneintritten,
die schnell und nach dem Alles oder Nichts-Prinzip verlaufen. Wie bereits
erwähnt, sind nicht alle Cdk-Formen für die Zellzyklusprogression essentiell.
Die Expressions- und Aktivitätsmuster der wichtigsten Zykline bzw. Cdk/Zyklin-Komplexe
sind zusammen mit weiteren, zum Teil nur phasisch aktiven Zellzyklusregulatoren
in der folgenden Abb. dargestellt.
Abb.:
Oszillationen zentraler Zellzyklusregulatoren Die Regulation des G1/S-ÜbergangsDie
Entscheidung zum Eintritt in die S-Phase wird in Vertebratenzellen über
den Phosphorylierungsstatus des Retinoblastomproteins pRb und seiner
verwandten Proteine p107 und p130 reguliert. Diese Inhibitoren
kontrollieren eine Reihe von Transkriptionsfaktoren, zu denen auch die
E2F-Familie gehört (Goodrich und Lee, 1993; Sellers und Kaelin, 1996).
Die mehrfache Phosphorylierung von pRb (Hyperphosphorylierung) in der späten
G1-Phase setzt E2F-1 und seine aktivierenden Bindungspartner DP1 oder 2
aus den pRb-Komplexen frei. Die freien E2F-1/DP-Heterodimere
transaktivieren eine große Zahl von Promotoren S-Phase-spezifischer Gene,
zu denen u.a. Enzyme der Nukleotidsynthese und Zellzyklusregulatoren wie
Zyklin E, Zyklin A, PCNA (proliferating cell nuclear antigen) und c-Myc gehören. Zusammen mit
der an Zyklin E gebundenen Cdk2, die an der Initiation der DNA-Synthese
beteiligt ist (Dutta und Bell, 1997; Grafi, 1998) induzieren sie den
Eintritt in die S-Phase (Bartek et al., 1997; Johnson und
Schneider-Broussard, 1998; Bateman, 1998). Neben den Zyklin-abhängigen
Kinasen sind vor allem durch extrazelluläre Wachstumsfaktoren stimulierte
Signalkaskaden u.a. über die Mitogen-aktivierte Proteinkinase (MAPK),
c-Src und c-Abl (zwei zytosolische Tyrosinkinasen) sowie die Januskinasen
(JAK) an der Regulation der pRb-Phosphorylierung und somit dem Eintritt in
die S-Phase beteiligt (Treisman, 1996; Roussel, 1998). Wie diese
Signalwege nach Serumstimulation in die Regulation der Cdk- und
E2F-1-Aktivierung eingreifen ist in der nächsten Abb. vereinfacht
dargestellt.
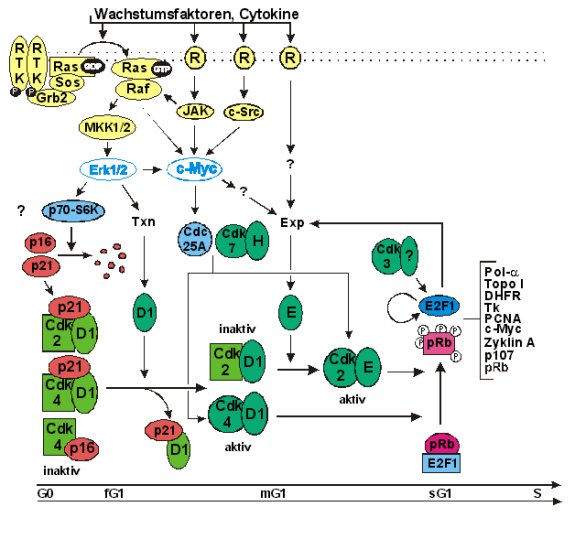
Abb.:
Modell der Regulation des G1/S-Übergangs
Zellen, die unter Serummangel kultiviert werden, enthalten inaktive Komplexe der Cdk2, 4 und 6 mit Zyklin D1 und p21 bzw. p16 (Serrano, 1997). Durch Wachstumsfaktoren werden die MAP-Kinasen Erk1 und 2 in der frühen G1-Phase aktiviert. Beide Kinasen hemmen die Expression der Cdk-Inhibitoren und induzieren Zyklin D1 (Lewis et al., 1998; Peeper und Bernards, 1997; Roussel, 1998). Diese Vorgänge verändern die Zusammensetzung der vorhandenen Cdk-Komplexe. Zyklin D1 verdrängt p16 kompetitiv und rekrutiert p21 in Komplexe, die keine Kinase enthalten (Sequestrierung). Hierdurch werden die Cdk4/D1-Komplexe in der mittleren G1-Phase aktiviert, während die Bindung von D1 an die Cdk2 inhibitorisch ist (Morgan, 1997). Erst die seruminduzierte Stabilisierung der Zyklin E-mRNA und die Zunahme des Zyklin E-Gehalts in der mittleren G1-Phase führen zur Aktivierung der Cdk2 (Oda et al., 1995; Othani et al., 1995). Die Cdk-aktivierende Kinase (CAK, Cdk7/H) wird in der frühen G1-Phase transkriptionell durch c-Myc induziert und von Raf-1 aktivierend phosphoryliert. Ihre Aktivität ist für die G1-Progression nicht limitierend (Facchini und Penn, 1998; Roussel, 1998).
Die
aufeinanderfolgenden Phosphorylierungen von pRb durch die Cdk4/D1- und
Cdk2/E-Komplexe setzen E2F-1 frei und induzieren eine erste Erhöhung des
E2F-1-Transaktivierungspotentials in der späten G1-Phase (Kitagawa et
al., 1996; Johnson und Schneider-Broussard, 1998; Lundberg und Weinberg,
1998). Wahrscheinlich folgt hierauf noch ein weiterer Aktivierungsschritt
der durch die Cdk3 katalysiert wird. Der G1-Bindungspartner dieser Cdk ist
bisher nicht bekannt; in der frühen S-Phase bindet die Cdk3 an Zyklin A (Morgan,
1997). Die aktivierten E2F-1-Komplexe verstärken das Signal zum Eintritt
in die S-Phase über mehrere positive Rückkopplungsschleifen indem die
Transkription der E2F-1-, Zyklin E- und A-Gene stimulieren. Dies führt zu
einer schnellen Zunahme der E2F-1-Aktivität und zum irreversiblen
Eintritt in die S-Phase (Johnson und Schneider-Broussard, 1998; Bateman,
1998). Die
G1-Progression proliferierender Zellen unterscheidet sich hauptsächlich
in Bezug auf Zyklin D1 und die Cdk-Inhibitoren von diesem Modell. Eine
dramatische Zunahme der Zyklin D1-Expression ist in proliferierenden
Zellen oft nicht nachweisbar. Da in diesen Zellen der Gesamtgehalt an
Kinaseinhibitoren ebenfalls deutlich geringer ist als in quieszenten
Zellen, kann eine ausreichende Cdk4 und 6-Aktivierung auch ohne deutliche
Expressionssteigerung von Zyklin D1 erreicht werden (Nigg, 1995; Morgan,
1997). Die Regulation des G2/M und M/G1-ÜbergangsIm
Verlauf Zellzyklusprogression von der G2-Phase bis zur Vollendung der
Mitose gibt es zwei Kontrollpunkte, die über komplexe Regulationswege mit
dem Aktivitätsstatus der Cdk1/B1 (MPF, mitosis
promoting factor) gekoppelt sind. Vom Ende der G2- bis in die
Prometaphase wirkt der sog. Kernhüllenabbaukontrollpunkt (NEB-Kontrollpunkt),
an dem Zellen z.B. durch UV-Strahlung arretiert werden. Kennzeichnend für
diesen Typ des Zellzyklusarrests sind ein hoher Zyklin B1-Gehalt, aber
eine aufgrund der inhibitorischen Phosphorylierung von Threonin 14 und
Tyrosin 15 nur geringe Cdk1/B1-Aktivität. Wird dieser Kontrollpunkt
aktiviert, so werden die Zellen in G2 arretiert bzw. Vorgänge der
Prophase rückgängig gemacht und die Zellen in einem G2-ähnlichen
Zustand arretiert. Nach der Aktivierung des MPF beginnen die Zellen mit
dem Kernhüllenabbau und sind durch Strahlung nicht mehr arretierbar (King
et al., 1994; Rieder und Khodjakov, 1997). Der
zweite Kontrollpunkt der M-Phase detektiert nicht Mikrotubuli-gebundene
Kinetochore und ist durch eine hohe MPF-Aktivität gekennzeichnet. Er wird
z.B. durch Mikrotubuli-depolymerisierende Agenzien aktiviert und liegt am
Ende der Prometaphase. Funktionell läßt sich dieser Kontrollpunkt in
zwei APC (anaphase promoting
complex, cyclosome)-gesteuerte Prozesse
unterteilen. Der APC ist ein Multienzymkomplex, der
Substraterkennungsfunktion für den Ubiquitin- und D-Box (destruction
box)-abhängigen Abbau über das 26S Proteasom hat (Pagano, 1997;
Pickard, 1997). Er wird durch Kinetochorproteine wie z.B. MAD2 gehemmt,
bis alle Kinetochore an Mikrotubuli gebunden sind. Nach der vollständigen
Ausbildung der Äquatorialplatte wird der APC in der Metaphase u.a. durch
den MPF aktiviert. Der Abbau mehrerer Anaphaseinhibitoren (u.a. der Säugerhomologe
von CUT2/PDS2 aus Hefe) ermöglicht die Trennung der Schwesterchromatiden
und die Anaphaseprogression. Nach einer Verzögerungsphase wird auch
Zyklin B1 durch den APC dem Abbau zugeführt, wodurch in der Telophase die
Vorgänge des NEB rückgängig gemacht werden und die Zytokinese einsetzt
(King et al., 1994, 1996; Pagano, 1997; Rieder und Khodjakov, 1997). An
der Regulation des MPF in der G2- und M-Phase sind u.a. Oszillationen der
MAPK-, PkA- und PkC-Aktivität beteiligt (Abb.). MAPK- und PkA-Aktivatoren
sowie TPA verhindern in G2-Zellen den Eintritt in die M-Phase durch
Aktivierung des Kernhüllenabbau-Kontrollpunkts. Die Erhöhung des
intrazellulären, freien Ca2+ dagegen ist ein positiver
Stimulus. Die Hemmung der MAPK und der PkA ist für den G2/M-Übergang
notwendig. Nach dem Eintritt in die M-Phase werden beide kurzfristig
aktiviert und vor der Zytokinese wieder abgeschaltet (King et al., 1994;
Walter et al., 1997).
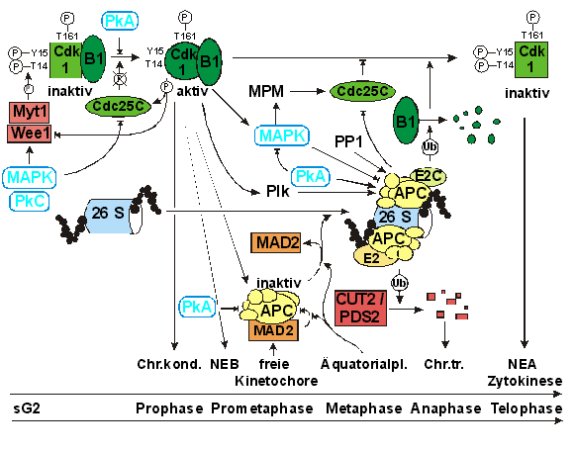
Abb.:
Modell der G2/M- und M/G1-Regulation 26S: 26 S Proteasom. APC:
anaphase promoting complex. B1:
Zyklin B1. Cdc25C: Phosphatase. Chr.kond.:
Chromosomenkondensation. Chr.trennung:
Chromatidentrennung. CUT2: cells
ultimately torn, Anaphaseinhibitor. E2:
Ubiquitin-konjugierendes Enzym. E2C:
Zyklin B-spezifisches E2. MAD2:
mitosis arrest deficient 2, Teil des Kinetochorkontrollpunkts. MAPK:
Mitogen-aktivierte Proteinkinasen Erk1, 2. MPM:
MPM-Kinasen. Myt1: Kinase. NEA: nuclear
envelop assembly. NEB: nuclear envelop breakdown. PDS2:
Anaphaseinhibitor. PkA:
Proteinkinase A. PkC:
Proteinkinase C. Plk: polo-like kinase. PP1:
Proteinphosphatase 1. Ub:
Ubiquitin. Wee1:
Kinase. ®: positiver Einfluß. ®|: hemmender Effekt.
|
|||||||||||||||||||||||||||
Copyright Information Portions of this work may be individually downloaded, copied, and cited for the personal and educational purposes of individuals and organizations in their work, provided that proper attribution and context are given. Commercial reproduction or multiple distribution of any kind is prohibited. All rights reserved. Reproduction in whole or in part in any form or medium without express written permission of the author is prohibited (kinetophil@yahoo.com). |